
Почвенные грибы – важный фактор распада, патогенеза и здоровья растений и почвы, включая круговорот питательных и органических веществ, плодородие почвы и стабильность агрегатов. В сельскохозяйственных почвах присутствует как минимум 25 000 видов грибов, это около 70% биомассы микроорганизмов. Рост грибов – функция наличия углерода. Длина гиф обычно колеблется от 3 до 300 м/г почвы. Большинство грибов живут в ризосфере, которая богата органическим углеродом, поступающим из протеинов, аминокислот, органических кислот и сахаров, выпускаемых корнями, мукополисахаридной слизью корней и отпавшими клетками корневых чехликов.
Вклад грибов в агроэкосистему трудно оценить из-за отсутствия точных методов измерения биомассы грибов и их деятельности. В агроэкосистемах выделяют три важные группы почвенных грибов: сапрофиты, патогены и симбионты. Основная часть биомассы грибов – это древовидные микоризные (ДМ) грибы, живущие в симбиозе.
Грибы-сапрофиты
Сапрофиты – основные редуценты растительных остатков, тогда как бактерии и специфические грибы расщепляют животный и микробный материал. Поскольку сапрофиты не слишком активны в роли редуцентов, ученые часто не обращают на них внимания, но жизнь на нашей планете не смогла бы продолжаться, если бы эти грибы не перерабатывали такие основные элементы, как С, N, Р и К. Тогда мы бы уже 100 раз были засыпаны неразложившимися листьями, корнями и другими растительными материалами. Несмотря на то что эти грибы играют очень важную роль в движении питательных веществ, они находятся в основном на растительных остатках, расположенных на поверхности почвы, и составляют менее 1% от общей микробной биомассы на глубине до 20 см.
В большинстве своем сапрофиты свойственны определенному виду растений, а типу субстрата. Субстраты можно разделить на несколько групп:
1) простые, растворимые углеводы;
2) нерастворимые углеводы;
3) лигнин и целлюлоза.
Грибы, использующие растворимые углеводы – в основном зигомицеты, с коротким жизненным циклом, характеризующимся быстрым ростом и образованием спор. Нерастворимые углеводы расщепляются в основном аскомицетами, которые присутствуют в почве повсеместно и зачастую производят антибиотики или вырабатывают устойчивость к ним, чтобы успешно конкурировать за субстраты. Грибы, расщепляющие лигнин и целлюлозу – медленно растущие базидиомицеты, обычно используют другие субстанции в качестве источника углеродной энергии, но при этом содержат энзимы, которые расщепляют лигнин и целлюлозу на субстраты, впоследствии перерабатывающиеся другими микроорганизмами.
Грибковые патогены растений
Патогены растений чрезвычайно важны для агроэкосистем, грибковые инфекции приносят огромные экономические убытки. Патогены обычно попадают в ткани растений через молодые части, корневые волоски или раны. Типичными примерами корневых патогенов являются Fusarium, Phytophtora, Pythium и Rhizoctonia.
У растений выработалось несколько механизмов защиты от грибов-патогенов. Физические барьеры, например, слизь на корнях растений и стенках клеток, – первая линия обороны. Другие защитные механизмы:
1) гиперчувствительная реакция (отмирание пораженной ткани вокруг точки заражения, позволяющее остановить распространение);
2) одревеснение стенок клеток;
3) синтез целлюлозы или каллозы;
4) накопление фитоалексина;
5) выделение гидролитических энзимов;
6) синтез ингибиторов протеиназы;
7) накопление гликопротеинов, богатых гидроксипралином.
Сельскохозяйственные методы, особенно методы самовосстанавливающегося земледелия, позволяют контролировать грибковые патогены за счет:
1) использования обработки почвы для заделки органов размножения подальше от новых корней;
2) увеличения количества культур в севообороте или посева буферных полос, а также посева других культур в междурядьях;
3) использования устойчивых покровных культур или пара для ограничения выживания органов размножения;
4) применения фунгицидов или методов биологического контроля, например, компостирование, использование микопаразитов, т.е. грибов, являющихся паразитами по отношению к другим грибам, или конкуренции микробов;
5) выращивания культур с защитными механизмами, в которых участвует множество генов, или ограничения количества культур, защищающихся лишь какими-то одними специфическими генами на поле.
Используя эти методы, можно сократить количество или вообще избавиться от патогенов, т.к., в отличие от сапрофитов и большинства симбионтов, патогены обычно свойственны определенному виду растений.
Биотрофные грибы-симбионты
При симбиозе как растение-хозяин, так и внедрившиеся грибы получают пользу, которая превышает связанные с этим сложности симбиоза. Грибам не хватает углерода, поэтому они устанавливают связи с растениями для получения углеродосодержащих продуктов фотосинтеза. Некоторые из этих грибов могут быть сапрофитами (например многие эктомикоризные или эндомикоризные виды после первого прорастания) или патогенами, но в большинстве своем симбиоз – нормальная форма сосуществования. Биомасса растения-хозяина увеличивается благодаря свободному получению питательных веществ, особенно таких малоподвижных, как Р и Zn.
Грибы-симбионты вызывают физиологические изменения в растении-хозяине. Когда из-за наличия грибов возникают изменения в химическом составе стенок клеток, увеличивается устойчивость к заболеваниям или изменяется реакция гиперчувствительности, позволяющая замедлить эти процессы или избавиться от инфекции вообще. Растения стимулируют собственную колонизацию грибами-симбионтами, увеличивая выделение из корней, ускоряющее прорастание спор и рост ростковой трубочки; повышая ветвление корней, увеличивая площадь колонизации, изменяя проницаемость мембраны клеток.
Древовидные микоризные грибы
Из четырех основных типов микоризных грибов эндомикоризные (ДМ) грибы представлены наиболее широко и повсеместно распространены в агроэкосистемах. ДМ грибы составляют 5-50% общей биомассы микробов и присутствуют на 70% сосудистых видов растений, т.е. практически на всех культурах. Исключением являются некоторые представители семейства крестоцветных, а именно брокколи, цветная капуста, крамбе и рапс. Традиционно крестоцветные считаются немикоризным семейством. Однако было отмечено, что микориза колонизирует около 33% представителей этого семейства.
Эндомикоризные гифы могут колонизировать до 80% длины корней растения-хозяина, проникая в стенки клеток и образовывая разветвленные структуры, называемые арбускулами, где происходит обмен питательными веществами и углеродом. Внутрикорневую колонизацию осуществляют гифы, споры, арбускулы. Колонизацию можно легко измерить и использовать как показатель активности грибов, но считается, что она составляет лишь небольшую часть биомассы. Внекорневые гифы и споры составляют около 80-90% биомассы ДМ грибов.
Около 12-30% фотосинтетического углерода растений перемещается в почву в форме углеводов, способствующих росту и развитию грибов. Эти углеводы быстро превращаются в углеводные спирты для поддержания движения углерода к грибам. Затраты углерода, которые несет растение, уравновешиваются увеличением доступа к большему объему почвы посредством гиф грибов.
У гиф соотношение площади поверхности к объему намного больше, чем у корневых волосков, поэтому они могут уходить на 8 см дальше зон, где нет питательных веществ вокруг корней (рис. 1).
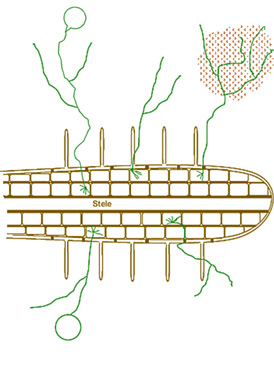
Рис. 1. Гифы древовидных микоризных грибов могут охватывать большее пространство почвы, чем корни и корневые волоски, и образуют рамку, вокруг которой формируются почвенные агрегаты
Это дает возможность ДМ грибам извлекать даже очень малоподвижные питательные вещества, например, фосфаты. Большие затраты углерода на вынос Р компенсируются повышением способности растения-хозяина проводить фотосинтез благодаря увеличению поверхности листьев и эффективности фотосинтеза. Микориза – наилучший механизм получения фосфора, особенно в условиях стресса.
Микоризные грибы могут приносить и другую пользу в разной степени, например, более эффективно впитывать азот и такие микроэлементы, как Fe, Cu и Zn, а также воду, подавлять заболевания, защищать от токсичных тяжелых металлов; улучшать структуру почвы. Микориза сокращает рост патогенов, особенно грибковых, увеличивая сопротивляемость растения-хозяина (вызывая ответную реакцию), изменяя химический состав корневых выделений, что вызывает рост микробов, являющихся антагонистами патогенам, увеличивая способность конкурировать за фотосинтетический углерод и сокращая количество пораженных зон. Как и другие преимущества микоризных связей, степень и направление влияния ДМ грибов на устойчивость к заболеваниям зависит от генотипа растения-хозяина, вида ДМ и изолята, времени колонизации ДМ, наличия других почвенных организмов и абиотических факторов.
Растения – хозяева микориз были обнаружены на большинстве участков, загрязненных тяжелыми металлами, но эти грибы обычно не изучались. В ходе экспериментального выращивания растений в горшках было обнаружено, что помимо микроэлементов микоризные грибы поглощают токсичные тяжелые металлы, например, Cd и Pb. Вынос металлов зависит от уровня плодородия почвы, концентрации металла, рН, растения-хозяина и вида ДМ и может мешать выносу Р растением.
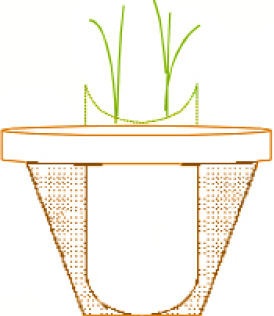
Одновидовые древовидные микоризные грибы можно вырастить на различных растениях-хозяевах для изучения накопления гломалина в стерильном песке или, как в данном случае, в песчано-угольной среде, где корни находятся внутри нейлоновой сетки, а гифы грибов, которые прорастают через ячейки сетки в окружающую среду, в отделение для гиф, при выращивании одного вида растений
Помимо благотворного влияния на здоровье растения гифы грибов улучшают структуру почвы, помогая образовывать нерастворимые в воде агрегаты. Микоризные грибы также улучшают здоровье ризосферы, стимулируя выделение из корней, которое способствует росту других почвенных организмов.
Гломалин – гликопротеин, производимый ДМ грибами
Выделение гломалина – гликопротеина, производимого ДМ грибами, – привело к переоценке вклада грибов в органическое вещество почвы (ОВП) и стабильность агрегатов. Гломалин был открыт учеными Министерства сельского хозяйства США в 1993 году во время работы по выделению моноклональных антител, которые реагировали бы с ДМ грибами.
Гломалин содержит высокие концентрации железа (2-12%), и недавно было установлено, что Al- и Fe-гидроксилы участвуют в формировании агрегатов, прикрепляя органическое вещество к частицам глины.
Характеристика гломалина
Гломалин, экстрагируемый из почвы, очень похож на тот, что экстрагируется из одновидовых горшочных культур. В образцах были выявлены незначительные различия элементарных составляющих гломалина, но анализы структурных групп (ЯМР, ГХ-СМ и КЭ) и SDS-PAGE показали, что гломалин, выделенный из почвы, аналогичен гломалину с гиф. Риллинг и его коллеги (Rilling et al., 2003), а также Стайнер и Риллинг (Steiner and Rilling,
2003) изучали разложение гломалина после инкубации почвы. Согласно их расчетам, период оборачиваемости гломалина – 6-42 года. Эти исследования инкубации говорят о том, что фракции прочного гломалина с длительным периодом существования характеризуются большим временем оборачиваемости.
Эксперименты по определению структурных единиц гломалина проводятся и сегодня. Информация, которой мы обладаем, говорит о том, что гломалин состоит из белковых, углеводных и алифатических (в основном полимеризированных) компонентов и связывает многовалентные катионы, т.е. Fe и Al. Протеиновый компонент составляет приблизительно 30-40% молекулярной структуры. Углеводный компонент составляет 3-6%. Алифатические группы составляют 20-70%. В гломалине содержится 2-12% железа.
Гломалин – основной компонент органического вещества почвы
Было проведено исследование, в котором сравнивались концентрации гломалина в гуминовой кислоте (ГК), фульвовой кислоте (ФК) и макрочастицах органического вещества (МОВ) на восьми необрабатываемых участках в США. Все фракции были названы по методам экстракции. Для извлечения каждой фракции был использован подходящий метод экстракции:
1) щелочная экстракция ГК и ФК, за которой следует кислотная сепарация;
2) экстракция гломалина цитратом;
3) разделение по плотности МОВ.
В ходе исследования было обнаружено, что гломалин представляет собой основную фракцию органического углерода почвы (ОУП; 22-27%), а выделяемая часть материала, которая раньше определялась как ГК и гумин, содержит гломалин.
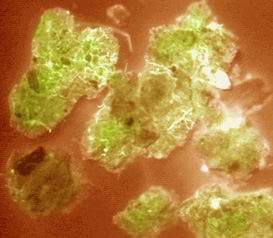
Гломалин и гифы древовидной микоризы на поверхности агрегата размером 1-2 мм, отделенного от почвы. Гломалин – белые пятна, подсвеченные благодаря иммунофлуоресцентному тесту, в котором используется моноклональное антитело против гломалина
Количества гломалина
Гломалин экстрагировался из почв различных экосистем США и мира с концентрациями ОГ,
колебавшимися от 2 до 14 мг/г почвы. Большие количества гломалина (ОГ) были обнаружены в необрабатываемых, вулканических почвах Японии и Гавайев (19 мг/г почвы и более 60 мг/г почвы соответственно) и в дубовом лесу на подзолистых почвах Ирландии (69 мг/г почвы).
Обычно в кислых почвах содержится больше гломалина, чем в известковых. В кислых почвах ниже уровень разложения и больше растворимых металлов (например, Fe и Al), которые могут способствовать увеличению концентраций гломалина, взаимодействуя с молекулами, замедляющими разложение. В залежных почвах уровень разложения ниже по сравнению с сельскохозяйственными, и больше ДМ грибов, т.к. в залежные почвы не поступает Р из удобрений и механическая обработка не разрушает сети гиф.
Концентрации гломалина изучались в нескольких сельскохозяйственных почвах с различными видами обработки, севооборота, удобрений, гербицидов и пестицидов. В большинстве этих систем была обнаружена корреляция между стабильностью агрегатов и концентрацией гломалина. При этом в системах No-till и минимальной обработки значения были самыми высокими. На экспериментальном участке в Мериленде на одном и том же поле изучались различные системы:
1) No-till, синтетические ресурсы (NT);
2) традиционная обработка, синтетические ресурсы (ТО);
3) минимальная обработка, органические ресурсы (МО).
Это позволяло определить, как управленческая система влияет на гломалин-С и МОВ-С. На участке с МО было наибольшее содержание гломалина-С, МОВ-С и максимальная стабильность агрегатов, тогда как при ТО и NT эти значения не отличались.
Гломалин и стабильность агрегатов
Потеря почвы в результате эрозии – серьезная проблема агроэкосистем. Пиментел и его коллеги (Pimentel et al., 1995) определили, что за последние 40 лет практически 1/3 обрабатываемых земель в мире была потеряна в результате эрозии, при этом скорость потери составляет 10 млн га/год.
Почвенные агрегаты важны для:
1) поддержания пористости почвы, что обеспечивает аэрацию и инфильтрацию, способствующие росту растений и микробов;
2) увеличения стабильности и защиты от ветровой и водной эрозии;
3) накопления углерода, защищая органическое вещество от разложения микробами.
Так как стабильность агрегатов и ОВП уменьшаются при обработке почвы, возможно, что ОВП (т.е. МОВ, гуминовые субстанции, молекулы, производимые микробами, и гифы грибов) принимает участие в формировании агрегатов, но точный механизм этого еще не до конца изучен. Формирование агрегатов – сложный процесс физических и химических взаимодействий.
При рассмотрении под электронным микроскопом можно увидеть, что агрегаты – это объединения почвенных минералов (частиц глины, мелкого песка и ила), небольшие остатки растений и микробов, бактерии, свободное аморфное органическое вещество и органическое вещество, тесно связанное с глиняным покрытием. Гифы грибов могут положить начало формированию агрегатов, обеспечивая рамку, на которой собирается органическое вещество. Затем химические процессы продолжают эту цепочку формирования и увеличения стабильности агрегатов, приклеивая к полисахаридам, покрывая водоотталкивающими полимерами, связывая минеральные частицы с органическими полимерами и объединяя органическое вещество и частицы глины при помощи поливалентных катионов. Намокание и высыхание, разбухание и сжатие глины, циклы замораживания-оттаивания, уплотнение и опутывание гифами грибов, а также мелкими корнями физически стабилизируют агрегаты.
Почвенные агрегаты могут быть разрушены дождем из-за возникновения чрезмерного увлажнения, дифференциального разбухания глины, механической дисперсии под действием кинетической энергии капель дождя и физиохимического распределения без защиты водоотталкивающего покрытия. Молекулы, участвующие в формировании агрегатов, увеличивают их устойчивость к действию воды и долгосрочную стабильность, потому что внутренние притягивающие силы этих молекул намного сильнее, чем внешние. При использовании систем минимальной обработки или No-till. Чейни и Свифт (Chaney and Swift, 1986) обнаружили, что стерня и мульча способствуют формированию агрегатов, т.к. при разложении органического вещества грибами возникают клеящие вещества, например, полисахариды и выделения корней.
Цезарь-ТонТрат и Кохран (Ceasar-TonThrat and Cochran, 2002) обнаружили, что лигнинолитические базидомицеты производят большие количества полисахаридов, гликолипидов или гликопротеинов, которые связываются с почвенными частицами и стабилизируют их в агрегатах, которые не разрушаются в воде. Однако многие из полисахаридов, произведенных в процессе разложения микроорганизмами, склеивают агрегаты быстро, но при этом растворяются в воде и являются недолговечными, следовательно, не продлевают срок стабильности агрегатов. Органическое вещество почвы, содержащее высокие концентрации алифатических групп, например ГК, могут увеличивать стабильность агрегатов и стабилизировать органические материалы на длительное время. Эти алифатические водоотталкивающие группы и полимеры являются основными веществами, повышающими стабильность агрегатов в воде. Они увеличивают угол касания для проникновения воды, в результате чего уменьшается уровень инфильтрации и насыщения, снижается степень промокания и усиливаются связи внутри агрегатов. Соединения между органическим веществом и аморфными соединениями Fe и Al также снижают уровень промокания агрегатов.
Гломалин приводит к стабилизации агрегатов, т.к. он отпадает с гиф на окружающее органическое вещество, связывается с глиной, что, возможно, происходит благодаря катионным связям с участием железа; и обладает водоотталкивающими характеристиками, которыми он обязан множеству алифатических групп. Об этом свидетельствуют результаты ряда экспериментов, в которых концентрация общего и особенно иммунореактивного гломалина положительно коррелирует с процентом нерастворимых в воде агрегатов как в сельскохозяйственных, так и в залежных землях.
Гломалин и стабильность агрегатов можно использовать как показатели для количественного определения изменений, происходящих в почве, при переходе от постоянной вспашки к No-till в течение короткого периода времени (1-3 года). Количество гломалина и стабильность агрегатов значительно увеличивались после 2 лет использования технологии No-till. На участке, где почва не обрабатывалась в течение 15 лет, концентрации гломалина (ОГ) и стабильность агрегатов были намного выше, чем на каком-либо другом. Это исследование говорит о том, что уровни гломалина, МОВ и стабильности агрегатов в системах минимальной обработки почвы продолжают увеличиваться с течением времени.
Гломалин в условиях повышенного уровня СО2
Было проведено несколько исследований, в ходе которых сравнивались концентрации гломалина по отношению к стабильности агрегатов в условиях повышенного уровня СО2. В системах залежных земель на севере Калифорнии концентрации ОГ и ИРОГ увеличивались с повышением концентрации СО2 и удлинением гиф на одном участке, а также стабильностью агрегатов в агрегатных фракциях 1-2 мм и 0,25-1 мм. Длительное воздействие увеличенных уровней атмосферного СО2 из естественных источников в Новой Зеландии привело к линейному повышению процента колонизации корней ДМ грибами, удлинению гиф в почве, увеличению ОГ, ЛЭГ в соответствии с повышением уровней СО2. На поле сорго стабильность агрегатов, длина гиф и ЛЭГ повышались с увеличением уровня СО2. Как на пастбищах, так и на поле сорго стабильность агрегатов коррелировала с концентрацией гломалина. Эти исследования показывают, что в условиях повышенного уровня СО2 фотосинтетический углерод перемещается в почву и гломалин может становиться «емкостью», улавливающей этот углерод.
Вклад почвенных грибов в органическое вещество
Несмотря на то что можно выявить сотни метров гиф, биомасса грибов обычно недооценивается, и в настоящий момент вклад грибов в ОВП по массе количественно не подсчитывается. Стивенсон (Stevenson, 1994) подсчитал, что количество грибов в почве составляет 10-20 млн/г, тогда как количество бактерий – более 1 млрд/г почвы. Олссон и его коллеги (Olsson et al., 1999) на основе анализа фосфолипидных жирных кислот определили, что сухой вес гиф ДМ грибов составляет 0,03-0,35 мг/г почвы. Одними из наиболее распространенных организмов в мире являются медленно растущие грибы почвы. Например, базидомицет Armillaria bulbosa был обнаружен в почве, занимающей 15 га, весящей более 10 000 кг, возраст которой составлял более 1500 лет (Paul and Clark, 1996).
Почвенные грибы влияют на формирование и функцию ОВП. При разложении грибами-сапрофитами элементы МОВ, например, азот и фосфор, превращаются из недоступных органических соединений в доступные неорганические источники питательных веществ. Разложившийся растительный материал затем становится частью гуминовой фракции почвы. Помимо этого ДМ грибы и гломалин помогают стабилизировать ОВП, участвуя в формировании агрегатов.
Вообще разнообразие почвенных организмов снижается в зависимости от методов ведения сельского хозяйства, а уровни минерализации увеличиваются, делая С питательным элементом, которого не хватает грибам. Биомасса грибов обычно положительно реагирует на No-till. Например, Фрей и его коллеги (Frey et al., 1999) обнаружили, что длина гиф грибов на поле, на котором применялась технология No-till, была в 2-2,5 раза больше, чем в системах традиционной обработки почвы. Грибы хорошо растут в системе No-till, потому что:
1). не разрушается сеть гиф;
2). грибы могут связываться с растительными остатками и использовать пространственно разбросанные питательные вещества, особенно С и N;
3). грибы могут оставаться активными даже в сухих зонах или в порах, заполненных воздухом.
После открытия гломалина и корреляции между иммунореактивной фракцией гломалина и стабильностью агрегатов этот гликопротеин оказался полезным в виде показателя качества почвы и микоризного влияния. По мере появления информации о структуре этой молекулы и ее различных массивах мы все лучше понимаем роль гломалина в агроэкосистемах. Из того, что уже известно, можно сделать вывод, что гломалин – основной компонент ОВП и важен для поддержания функционирования агроэкосистем.
Управление почвенными грибами
Для увеличения количества органического вещества самовосстанавливающаяся агроэкосистема – та, в которой внутренние механизмы и ресурсы могут поддерживать продуктивность, быстро восстанавливаться после разрушения (например механической обработки) и удерживать вредителей и болезни на допустимых уровнях при минимальном воздействии извне. В сельскохозяйственных почвах содержатся неестественно высокие количества Р, N и К из удобрений, эти почвы разрушаются под воздействием механической обработки и зачастую покрыты растительностью одного или двух видов растений (Gliessman, 2000; Muramoto et al., 2000). Чтобы уменьшить количество патогенов и увеличить биомассу, разнообразие и стимулировать функционирование грибов-симбионтов, можно внедрить одно или несколько управленческих решений из тех, что перечислены ниже (Carlile and Watkinson,
1996; Horwath and Paul, 1994; Stenberg, 1999; Wright and Anderson, 2000):
1). снизить количество вносимых удобрений (особенно удобрений с высоким содержанием Р), что увеличит способность симбионтов извлекать нерастворимые питательные вещества;
2). использовать системы почвозащитного земледелия или No-till, которые не будут разрушать сеть гиф;
3). увеличить количество культур в севообороте, производить посев культур в междурядьях или использовать буферные полосы либо лесополосы, что увеличит надземное разнообразие и уменьшит количество хозяев грибов-патогенов;
4). засевать покровные культуры вместо пара, что обеспечит присутствие живых корней, которые будут хозяевами грибов-симбионтов;
5). применять методы применять биологического контроля сорняков и вредителей, сокращая потери полезных грибов, возникающих в результате действия фунгицидов и других пестицидов.
Эти стратегии можно использовать в агроэкосистемах как способ управления сапрофитами, симбионтами и патогенами в почве, получая увеличение урожайности культур при минимальных затратах.
Кристин Николс и Сара Райт
Читайте также:




 2026-06-10
2026-06-10

